The sandmat group (sect. Anisophyllum) of Euphorbia is quite different from the other groups. It is likely the most distinctive and has commonly been referred to by its own genus (most often and most recently Chamaesyce). It is probably most commonly known in North America for spotted spurge. I have written fairly extensively and technically about the differences, but I'm going to attempt to simplify it all here. If you are interested in the more technical explanation, you might try reading the morphology section of my thesis.

Spotted spurge (Euphorbia maculata).
C4 photosynthesis and Kranz anatomy
Perhaps the most consistant and interesting charactaristic is the presence of C4 photosynthesis, which occurs in all but subsect. Acutae (i.e., Euphorbia acuta, E. angusta, and E. johnstonii). Of course, you can't really see C4 photosynthesis, but there is a cool trick you can use to tell if a plant has C4 or C3 photosynthesis. C4 photosynthesis comes with specialized anatomy called Kranz anatomy where the primary photosynthesizing cells are clustered around the veins. This makes the veins look darker than the surrounding tissue creating an interesting pattern.


Kranz anatomy in Euphorbia prostrata (left). Euphorbia angusta, the only species in section Anisophyllum confirmed to have C3 photosynthesis (right).
Dorsi-ventral (sided) stems and unequal leaves
In most species, there is a clear upper and lower side to the stems (i.e., a sidedness; also known as dorsi-ventral because the stems have a dorsal and ventral side). This is most obvious in prostrate species but is present in most upright species too. Take for instance hyssop spurge (E. hyssopifolia), nodding spurge (E. nutans), or graceful spurge (E. hypericifolia). In all these cases, the stems are generally quite upright, but the stems bend toward the tips so that the upper sides of the branches face the sun (usually described as arcuate ascending). This sided appearance comes in part from the asymmetric nature of the leaves. One side of the leaf (as divided by the midvein) is almost always longer than the other side. This makes the leaves more efficient when they turn sideways and allows the lobe of the longer side (what I have sometimes referred to as the cordate lobe) to overlap the stem, but the other side not to be obscured by the stem. The plants are so well evolved for a prostrate lifestyle that their leaves are built to make the most of it. There are rare exceptions to this rule (look at E. acuta for example), but it is a quick and pretty reliable technique to distinguish from the other groups.


Arcuate ascending habit and dorsi-ventral positioning in E. hypericifolia (left). Greatly exagerated assymetric leaves in a E. glyptosperma seedling (right).
Non-glandular stipules
Stipules are structures made of leaf tissue that grow on either side of a petiole. In some species outside Euphorbia, these can become leafy, thorny, or even twining (some tendrils are modified stipules). In Euphorbia they are almost always small, inconspicuous structures or may be absent altogether. In sect. Anisophyllum, they are generally small and white and can be found between the pairs of leaves. Each leaf produces one on each side of the stem (one on the dorsal, one on the ventral). Each individual stipule may become lobed to the point of appearing like more than one structure or can fuse with the other leaf's stipule on its side of the stem to form a scale. Though this can be useful for ID within sect. Anisophyllum, the key is that they are almost always present and almost never glandular. This is what helps to distinguish them from other groups.


Laciniate stipules of Euphorbia glyptosperma (left). Stipules of E. albomarginata that are fused together into a single white scale (right).
Sympodial branching (pseudo-dichotomous)
The branching is also interesting in sect. Anisophyllum. Within the group, each branch produced terminates (either stops growing, aborts, or is terminated by a cyathium) and is followed up by a pair of branches. This looks very similar to dichotomous branching which produces the same forking at the nodes, but this is slightly different. In true dichotomous branching (which can be seen in the wisk fern, genus Psilotum), the apical bud itself splits into two equal buds. In sympodial branching, the apical meristem itself stops and branches take over in its sted. This has led to the term pseudo-dichotomous branching (i.e., false dichotomous branching).
For distinguishing from other members of Euphorbia, the key is that members of sect. Anisophyllum have this branching right down to the base. Its very first apical meristem produces one pair of leaves and produces nothing else. The branches take over from there. In other Euphorbia groups, the inflorescences are often sympodial, but the vegitative stems are not. This has led to speculation that members of sect. Anisophyllum are the result of reduction of plants to only the inflorescence but this explaination as it has been traditionally proposed has faced some critisism, so I will not go into the details here. If anyone wants to learn more, I recommend Hayden (1988): Ontogeny of the Cotyledonary Region of Chamaesyce maculata (Euphorbiaceae). Regardless, this is another useful characteristic!


Sympodial branching and equal length in paired branches of E. golondrina (left). Seedling of E. glyptosperma showing blank, flat spot where the apical meristem would be in most other plants (right).
Ecarunculate seeds
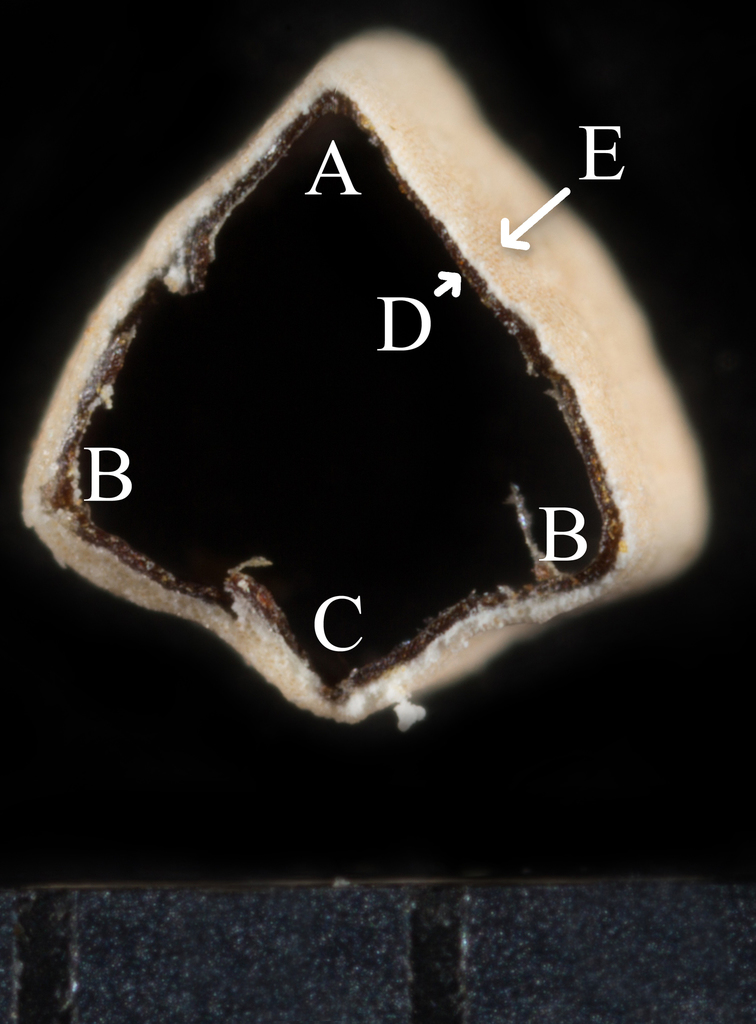
Lastly, members of sect. Anisophyllum have seeds that aren't carunculate. A caruncle is an outgrowth at the part of the seed that attaches to the fruit called the hilum. Many members of Euphorbia have carunculate seeds, but not members of sect. Anisophyllum. The closest they come is in the species E. carunculata but despite the name, the enlarged structure is an outgrowth of the micropyle (the part of the seed where the pollen grew through), not the hilum. For this one, it's probably easier to explain with pictures than words. If you want the full explanation or to know how seeds can be used in identification, I recommend reading my advanced seed morphology explanation.


Ecarunculate seed of E. lata ("E" represents the hilum), sect. Anisophyllum (left). Carunculate seed of E. tetrapora, subgenus Esula (right).

Outgrowth of the micropyle in E. carunculata, sect. Anisophyllum (left).
Taxonomically significant characteristics
These are characteristics you're likely to encounter when trying to key out members of this section.
Habit
There are several major forms. In the US, only the Hawaiian endemics, E. jaegeri, and E. chaetocalyx var. triligulata are woody (and they very from shrubs to trees). Euphorbia jaegeri, and E. chaetocalyx var. triligulata are subshruby or barely so while the Hawaiian endemics are much larger (some becoming trees). With herbaceous species, some have woody rootstocks, while some have non-woody rootstocks. This characteristic is significant outside the southernmost parts of the southern US deserts for differentiating perennials from annuals, but less so nearer the southern boarder. Among annuals, there are two main forms with intermediate species filling in the gaps between them. One is exemplified by the prostrate form like E. maculata or E. prostrata. The other is robust and upright and best exemplified in E. hypericifolia, E. hyssopifolia, and E. nutans.


Much branched growth from woody rootstock of E. fendleri (left). Typical annual growth of E. golondrina (center). Robust upright growth of E. hyssopifolia (right).
Inflorescences
Cyathia (flower-like inflorescences; more information here) may be densely clustered or solitary at the stems. Also, they may be congested along the stems or compacted into a round structure (i.e., a glomerule). There are gradations between all these characteristics but most keys try to distinguish between glomerules and other forms typically termed solitary.



Solitary cyathia of E. acuta (left). Conjested cyathia in elongated, leafy inflorescences of E. stictospora (center). Glomerules of E. hypericifolia (right).
One useful comparison to make when determining whether cyathia are solitary or in glomerules is to compare E. hyssopifolia and E. hypericifolia along with typical solitary cyathia. The glomerules are ultimately formed by shortining of the internodes after cyathial production and an eventual elimation of leaf production. An extra step of stems branching in the dorsal or ventral direction to the main axis to form a three-dimensional glomerule is sometimes taken in larger inflorescences. This can be contrasted with the two-dimensional plane in which all cyathia occur when cyathia are solitary.

Solitary cyathia of E. acuta (left). Inflorescences of E. hyssopifolia (center). Glomerules of E. hypericifolia (right).
The differences between the two congested inflorescence types are simply a matter of branching. In sect. Anisophyllum, branching is sympodial (pseudo-dichotomous) as mentioned above. This means that each branch forked into two (though one may be dominant over the other). In the glomerules, the forked branches are of essentially equal lengths. In the other dense inflorescence type, it appears that one branch is dominant over the other leading to what looks like a central axis with a proliferation of cyathia. In the case of inflorescences like E. hyssopifolia, the main distinction is in whether the inflorescence has leaves or not, or at least if it appears to. What the research is typically referring to is whether there are several leaves for the number of cyathia or few to none. As with many characteristics, it's rarely an absolute thing, but makes sense and is quite characteristic if you look at several examples.
Stem and fruit hairs
Depending on group, stem and fruit hairs may be completely unreliable as taxonomic characteristics or among the most reliable. In the US, the former case is rare and the latter is generally true. Examples where the presence or absence of hairs are unreliable include: Euphorbia polycarpa (all varieties), E. micromera, and California populations of E. serpillifolia. These plants are mostly found in the Sonoran Desert, Mojave Desert, and adjacent areas (the absence of hairs of E. micromera in Texas and New Mexico is actually a stable characteristic). Some examples where hairs may be present or absent outside of this area can be found in E. hyssopifolia, E. villifera, and sometimes E. abramsiana (also, potentially E. serrula in Mexican populations). Presence or absence of hairs is most often used, but sometimes the type of hairs is also important. For instance, when hairs are present in E. hyssopifolia, they are pilose (straight and longer) while the hairs of E. nutans are very short and crisped. Another common example: E. maculata has appressed fruit hairs while those of E. prostrata are pilose (except in rare cases where excessive sun exposure and/or trampling are involved).



Dust-like curly hairs of E. nutans (left). Hair stems and glabrous fruits of E. serrula (center). Hairy stems, fruits, and leaves of E. maculata (right).
Section in progress
Glands and appendages
Fruit shape
Styles